Two very different biology Citizen Science projects lead to insights into students´ and teachers´ learning. But learning must not be restricted to learning at school, a social learning theory widens the scope, where learning can take place. Join us as Martin Scheuch from University College for Agrarian and Environmental Pedagogy, Austria gives a seminar entitled Citizen Science and Learning Biology – examples and insights from two Austrian projects. Time & Date: 13:15 CET, 31.03.2023. Venue: 5F322 or over zoom through https://kau-se.zoom.us/my/magnuslovenwallerius.
On 21 March, Katharina Lapin will be giving a seminar with the title Management of invasive species in riparian forests to our department. Her work centers on the recognition and handling of invasive species in forested areas, but even their effects on forest ecological functions.
Katharina Lapin
As researcher at and head of the Forest Biodiversity & Nature Conservation department of the Austrian Research Centre for Forests (BFW), Katharina works on the interface of research and management of natural resources. This results in both scientific papers and handbooks that can be immediately applied in the management of Austrian forests. Read about the Centre for Forestry on https://www.bfw.gv.at/ (in German) and join us on Tuesday 21 March at 13:15 CET via <https://kau-se.zoom.us/my/magnuslovenwallerius>.
This post covers chapters 4 & 5 from Futuyma & Kirkpatrick’s book on Evolution (2018). The author of this post is Sam Shry.
We are all special: mutation and variation
It is quite obvious to see variation among individuals, but how does this variation come about? Chapter 4 takes us on a journey through the evolutionary processes of mutations, variations, and how they are linked to inheritance.
We start off small with DNA (or RNA for some organisms), genetic material made up of base pairs that are the building blocks of all forms of life. DNA is carried by chromosomes, with one inherited from each parent in humans. Interestingly enough, the complexity of an individual has no correlation with the size of their genome (e.g. salamanders are more complex gnomically than humans). These chromosomes contain genes, which are the areas of a chromosome that perform a function. The chapter covers in-depth the processes in which a gene’s DNA is used to make proteins, via transcription, splicing, and translation, but that’s all a little dense for a blog. What is interesting to me is how only 2% of our genetic makeup is actually used for making proteins while the rest is just there as non-coding “stuff”. Even in our DNA we are hoarders…We find, however, that protein synthesis is the driver that creates variation in our appearance (phenotype) and genetic makeup (genotype) via inheritance. Inheritance is really just a locus or gene that is passed down to the next generation, but this locus varies in its DNA sequence, giving rise to alleles, a term given to locus variation in a population. The mixing of genes in sexually reproducing organisms causes variation in allele frequencies for a population. This mixing is done through segregation and recombination, where segregation selects one or two gene copies from a locus of each parent and recombination combines gene copies from each parent. Segregation is interesting in how the frequencies of genotypes and alleles change over time. We have developed a mathematical model (Hardy-Weinberg equilibrium) in order to understand how evolutionary forces affect populations. Similarly, with recombination, we have developed a model (linkage equilibrium) in order to study the effects of recombination rates in loci. Of course, the models do not represent reality, but give us a starting point that we can use to examine how evolution is occurring in a population and what factors are having the largest effects on which loci. We also have a large proportion of species reproducing asexually, using horizontal gene transfer to move DNA between individuals, which is particularly important to bacteria’s ability to evolve antibiotic resistance.
Even with all this inheritable mixing, most variation occurs from mutations. Mutations are errors, messed up DNA replication that is the ultimate source of genetic variation. See! It’s good to mess up sometimes! They can be small, dented my car screw-ups like point mutations, which only occur at a single DNA base, or they can be hot mess situations like whole genome duplications tetraploidy, where offspring can’t reproduce with their parental population, resulting in a whole new species with a single mutation. Each type of mutation can occur at a different rate and can cause different degrees of effect. Usually, there is a linear trend in genome size and mutation rate, which can affect almost every aspect of an organism. Mutations can affect both an individual’s traits as well as their fitness. Natural selection tries to drive mutation rates down, but ironically it is mutations that drive adaption and allow us to survive. We’ve been hyping up mutations, but most of them are bad, with natural selection “picking out” what mutations are favorable in an individual’s environment, or in other words, what works in an environment doesn’t die or at least has a chance to “get-it on” before dying. There are also a vast number of fields that delve into the inheritable, but non-genetic, cultural changes that can alter the physical behavior and learning of the offspring, but we steer clear of that in this book.
Just natural selection bro
In chapter 5 we can explain the genetic theory of natural selection as fundamental and simple, though Darwin may disagree as he wrecked himself trying to understand this “simple” concept till he died. It wasn’t till after his death that Mendelian genetics was fused with Darwinian selection theory. Of all organisms, the peppered moth is our shining example of documented evolution via natural selection, ironically adapting to survive our coal burning in the industrial revolution. After documenting this discovery we went straight to work figuring out how we could exploit this beautiful, natural process by developing the technique of industrial artificial selection; selecting the best traits of what we consume to make it taste, tastier.
What is key to evolution by natural selection is that the trait (phenotype) has to be good enough to allow individuals to make lots of offspring (absolute fitness) and that the trait is inherited by the offspring, which in turn causes the evolution of the species. Fitness is a combination of the probability of survival to maturity and the expected number of offspring the individual will have. If we think of a male bodybuilder, he may seem “fit” physically, but from an evolution standpoint his fitness is only measured in his survival to maturity and the number of offspring he produces, which could be a problem due to all the steroids…
If an allele has higher fitness after a mutation, this will cause a positive selection of that mutation in the population. One example of this is the adaption to drinking cow’s milk (lactase persistence) in Northern European populations. We can calculate a measurable selection strength score for these beneficial alleles (selection coefficient), which can then be compared with the genetic variation in the population to predict the rate of adaptation. The rate of adaptation can vary based on the organisms’ generation time (bacteria vs. humans) and the allele dominance (dominant, non-dominant, recessive). There can also be crappy mutations (deleterious) that decrease fitness and unfortunately due to their lousy recessive qualities and low frequency, selection can’t remove them easily. Just like gambling, mutations are subject to chance and can be removed from a population even if they are beneficial, such as via genetic drift.
There is also a mix of situations where natural selection has side effects, some of which are good and others not so good. Some of these genetic side effects are genetic correlations, allele hitchhiking, and trade-offs between alleles, all of which are a by-product of selection that can have serious effects on the population. For example, the Scottish soay sheep have an evolutionary trade-off in their population where a polymorphism at a single locus causes homozygous individuals to grow vestigial horns. These sad-looking horns make it harder with the ladies (decreasing mating success) but increase survival (no macho fights).
Just as natural selection can lead to new forms of variation, it can also preserve variation available in a population via balancing selection. This can occur when hard-core heterozygote alleles have higher fitness than the homozygotes (overdominance) and lead to polymorphic equilibrium, where both alleles are maintained in the population. Balancing selection can also occur with frequency–dependent selection, where the frequency of the alleles determines their fitness. Specialization can also balance polymorphism, either via niche specialization or space specialization. Just as yin and yang, genetic variation can also be destroyed by natural selection via under-dominance and positive-frequency-dependent selection. Almost nothing can escape time though, with alleles spreading to fixation in the population, with the deep, thought-provoking idea that “the outcome of evolution is determined by where the population begins” (page 125).
Where a population begins is important, because natural selection is continuously adapting species to the environment in which they live, making this coupling between species to the environment. Fitness can increase or decrease based on the organism’s mutations, but the same ebbs and flows in fitness can be the result of environmental changes too.
To measure the fitness of a population over time, mean fitness can be calculated and compared between populations. This theorem of natural selection mathematically demonstrates that populations evolve via natural selection in order to increase their average survival and reproduction through time. The measurable adaptive landscape is the balance between fitness selection, environmental gains, mutations, etc., and though the fundamental theorem doesn’t exactly apply to real-world situations sometimes (typical math), it acts as a guide to understanding approximately what the hell is going on in the population from an evolutionary point of view. Some instances where the math is way off is when selection is frequency dependent, which can lead to mean fitness decline of the population. The same can be said for competition within a population, where some individuals develop nasty traits that become dominant in the population, but then turn out to bring down the fitness of the group.
Most of the time though, these mutations, usually bad, are killed off (deleterious). Selection does an adequate job of “purifying” our DNA from these crap mutations, but the problem is they can reappear at almost the same rate, causing the numbers to balance out. What’s interesting is it doesn’t matter how crap the mutation is (strong or weak deleterious mutations), the population’s mean fitness still decreases by the same amount. It’s important to understand how this load of crap (mutation load) is spread throughout the genome in order to understand its effect on the mean population fitness. These deleterious mutations cause a lot of problems in humans a.k.a. death, but modern medicine has decreased and postponed these deaths, and individuals have lived long enough to reproduce, making a controversial discussion as to the future impact this will have on humans, but maybe concentrating on climate change and keeping us from catching on fire is a more pressing issue
Jan Thiele from Thünen Institute for Biodiversity, Braunschweig-Germany will be giving a seminar today 7th March 2023 at 13.15 CET live over zoom. The title of this seminar is Effects of historical land-use and landscape structure on arthropod and plant communities in grasslands. To attend visit https://kau-se.zoom.us/my/magnuslovenwallerius
This is a literature course on the book Evolution by Douglas J. Futuyma & Mark Kirkpatrick (Fourth Edition) during the first half of 2023. This write-up covers chapters 1 to 3 and is authored by Louis Addo (a Ph.D. student at KAU)
The historical background of Evolution Biology
Evolution in biology refers to the shift in the heritable traits of biological populations over successive generations. Charles Robert Darwin (English naturalist and biologist, Figure 1. left) and Alfred Russel Wallace (an English naturalist, explorer, geographer, anthropologist, biologist and illustrator, Figure 1. right) separately developed the theory of evolution by natural selection in the middle of the 19th century, and it was extensively outlined in Darwin’s book On the Origin of Species. The hypotheses of evolution postulated that all living things share a common ancestor, and that natural selection acting on genetic variations is responsible for the changes in their populations over time. The theory of evolution became a scientific fact supported by other scientists in paleontology, genetics, and biochemistry. Prior to the postulation of the theory of evolution, the dominant worldview was that each species was uniquely created by God and had set characteristics. However, this worldview was challenged by the Enlightenment movement leading to the emergence of science. The foundation for evolutionary thought was laid by astronomers and geologists who postulated theories about the creation of stars, planets, and the earth including the changes the earth has gone through as well as its many extinct species. Unique to Darwin’s theory were its five components explaining concepts “descent with modification” and “natural selection”.
Figure 1 Famous fathers of evolution: Charles Robert Darwin (left) and Alfred Russel Wallace (right) (Futuyma & Kirkpatrick pp 11-13)
The five distinct components of Darwin’s theory of evolution are natural selection which means that species characteristics change over time to meet changes in their environment, evolution which proposes that organisms change over time, gradualism which means that the differences between even fundamentally different organisms have evolved through intermediate forms rather than by large leaps, common descent which suggests that species diverged from shared progenitors and that species might be seen as one enormous family tree that represents ancestry, and finally population change which means that evolution occurs by changes in the proportions (frequencies) of different variant kinds of individuals within a population.
Most scientists acknowledged the historical truth of evolution through descent with modification from common ancestors by the 1870s, but natural selection, which drives evolution, was not widely accepted until around 60 years after The Origin of Species was published. Many ideas, including neo-Lamarckian, orthogenetic, and mutationist theories, were put out during this period.
In general, the idea of evolution is a scientific truth that explains how ancestors give rise to various descendants and how species change over time. Biologists generally agree with the theory, and while work is still being done in some areas, the fundamental ideas of evolutionary theory are well-supported.
The “Tree of Life” and phylogenetics
The concept of the Tree of Life was first proposed in Charles Darwin’s book On the Origin of Species and proposes that all species, alive and extinct, descended from a single ancestral form of life that existed billions of years ago. The history of the events by which species or other taxa have arisen from common ancestors is called phylogeny, which is often represented by a phylogenetic tree that shows the genealogical relationships among the taxa. Anagenesis, which is the evolutionary modification of a lineage’s (species’) physical characteristics, and cladogenesis, which is the division of a lineage into two or more descendant lineages, are the two main processes that contribute to the development of higher taxa. After cladogenesis, anagenesis causes each descendant lineage to diverge even farther from the others. The branching order and the length of the branch in the phylogenetic tree can show which species are more closely and less distantly related to one another.
Phylogenetic studies reveal the evolutionary relationships between organisms and gene sequences, often showing the pattern of separate and divergent lineages. However, sometimes branches of a phylogenetic tree rejoin, forming a network rather than just a branching tree. Hybrid speciation is one example of this, which is especially common in plants, where some species evolve from hybrid crosses between two different ancestors. Another example is horizontal gene transfer (HGT) where genes are passed among organisms, enabling them to adapt to changing circumstances.
Phylogenetic analysis is an effective method for determining how different traits in organisms have evolved through time. This approach shows that species have evolved from similar creatures since it is based on homologous traits that have descended from them. The majority of an organism’s traits are changed from traits that already existed in its ancestors and do not develop independently. Although they often have comparable genetic and developmental bases, homologous physical characteristics between species can vary more than the finished products. For instance, in the 1970s, scientists analyzed the protein amino acid sequences of pairs of animals that split from their common progenitors at various points in time. They created a molecular clock by plotting the matching DNA sequence discrepancies against predicted divergence periods and finding that the rise in differences increased linearly with time (Futuyma & Kirkpatrick 41). Assuming the genes in these lineages had developed at the same pace as those in the animals with fossil records, this permitted calculation of the period of divergence even for lineages lacking a fossil record. Yet, there is no one molecular clock and rates of sequence evolution vary among different types of organisms.
The analysis of the evolution of different organismal traits with phylogenetic analysis offers overwhelming proof of evolution. It is feasible to evaluate homologous traits that are descended from common ancestors since characteristics of organisms nearly invariably evolve from pre-existing aspects of their predecessors. However, a character may be shared by several species but not necessarily in the same character state. It can be challenging to determine if two species’ traits are homologous, but embryological research and anatomical correspondence of position and structure, are the two most often used methods for making this determination are often successful methods.
The concepts of Natural Selection and Adaptation
Changes in the environmental adaptation requirements of organisms trigger biological changes in their behavior and physical features to ensure their existence in the altered environment. Thus, altered environments can cause natural selection on the genetic variation in many characteristics of a species or population. For instance, soapberry beetles adapted to new food sources by changing the length of their beaks, and several insect and plant species have developed a resistance to heavy metals and chemical pesticides. Fish overfishing has also affected behavior and accelerated the onset of sexual maturity. The process by which individuals with advantageous traits have a better chance of surviving and reproducing than those without, resulting in the preservation of preferred variations and the rejection of harmful ones, is referred to as natural selection. Darwin first introduced this idea in “On the Origin of Species.” When various biological entities consistently vary in fitness, natural selection takes place. It is important to understand that evolution, which may also be brought about by other mechanisms like genetic drift, is not the same as natural selection. The environmental conditions that force natural selection on a species are determined by its traits. Certain species can create ecological niches for themselves by eliminating elements of their environment so that these no longer force natural selection. Natural selection can take place at several scales, including genes, cell types, individual organisms, populations, and species.
Selfish genes, which multiply in the genome whether they are advantageous to the organism or not, are an illustration of gene-level selection. Gene-level selection can conflict with individual selection and cause harm to organisms. Selection among individuals occurs at a higher level than selection among genes. Characteristics develop through individual selection; altruism can lower individual fitness but may develop through social selection; cooperation develops through kin selection. By varying the percentage of species throughout time, species selection modifies the diversity of biological traits. It has an impact on organism disparity but not adaptations. For instance, more asexual populations experience extinction than sexual groups.
Adaptation in biology refers to both the process by which organisms evolve over generations to improve survival and reproduction, and to a characteristic that evolved by natural selection. A trait must be developed and give greater fitness than the ancestral condition in order to qualify as an adaptation. A feature that unintentionally fulfills a new purpose is known as a preadaptation. For instance, parrots can eat fruits and seeds with the help of their strong, pointed beaks but, in case a new resource is presented, they can also use it to feed on that resource. Exaptation is the process of adapting a characteristic for use unrelated to the one for which it was originally selected. In birds that initially evolved feathers to keep warm, an example of an adaptation would be the use of feathers for mating displays or flying. Pre-adaptation is another name for an exaptation. Exaptations and preadaptations are common in the early stages of the evolution of new adaptations. To determine if a certain characteristic is an actual adaptation, scientists examine the available data. Species traits are not always adaptations or random characteristics but can be flawed and limited, such as in the case of mammals that are unable to evolve beneficial modifications in the number of vertebrae. The vast diversity of life is the result of natural selection, as varied habitats and other factors may force selection on different traits within a species.
The next blog will cover chapters 4 and 5 and will be authored by Samuel Shry.
References
Futuyma, D. J., & Kirkpatrick, M. (2017). Evolutionary. Evolution (Fourth ed.). pp. 3-76. Sunderland, Massachusetts: Sinauer Associates, Inc.
Forskargruppen Naturresurs rinnande vatten (NRRV) har fått två ny forskningsprojekt beviljade. Lutz Eckstein är projektledare i ett projekt om effekter av korttidsreglering i vattendrag under vintern på ekologisk status av strandzonen som finansieras genom Kompetenscentret Svenskt vattenkraftcentrum (SVC) med 3,8 Mkr. Projektet är ett samarbete mellan Karlstads universitet (Eva Bergman, Larry Greenberg, Johan Watz) och Umeå universitet (Roland Jansson, Birgitta Malm-Renöfält) och kommer att undersöka vattendrag i både norra och södra Sverige. John Piccolo är projektledare i det andra projektet som kommer att fokusera på att upprätthålla vattenkraftsproduktion och värdefulla fiskpopulationer genom att utveckla Individual-Based Models (IBM) för att bedöma hur fiskpopulationer kan återställas samtidigt som flödesförhållanden för vattenkraftproduktion upprätthålls. Projektet finansieras genom Energimyndighetens program hållbar svensk vattenkraft (HåVa) med 2,9 Mkr och är ett samarbete mellan Karlstads universitet (Mahboobeh Hajiesmaeili och Johan Watz), Vattenfall AB (David Aldvén och Patrik Andreasson) och Fortum AB (Marco Blixt och Markku Lahti) och kommer att undersöka två viktiga vattenkraftsproducerande vattendrag i Sverige: Gullspångsävlen och Luleälven.
The River Ecology and Management research group (RivEM) has been granted two new research projects. Lutz Eckstein is the project manager in a project, which is financed through the Competence Center Swedish Hydropower Center (SVC) with 3.8 million SEK, on the effects of short-term regulation of rivers during winter on the ecological status of the riparian zone. The project is a collaboration between Karlstad University (Eva Bergman, Larry Greenberg, Johan Watz) and Umeå University (Roland Jansson, Birgitta Malm-Renöfält) and will investigate rivers in both northern and southern Sweden. John Piccolo is the project manager in the second project that will focus on sustaining hydropower production and high-value fish populations by developing Individual-Based Models (IBM) to assess how fish populations can be restored while maintaining streamflows for hydropower production, which is financed through the hållbar svensk vattenkraft (HåVa) program of Energimyndigheten with 2.9 million SEK. The project is a collaboration between Karlstad University (Mahboobeh Hajiesmaeili and Johan Watz), Vattenfall AB (David Aldvén and Patrik Andreasson) and Fortum AB (Marco Blixt and Markku Lahti) and will investigate two key hydropower producing rivers in Sweden: Gullspångsävlen and Luleälven.
Juliana Daniela-Ferreira (a postdoc researcher from SLU Uppsala) will be giving a talk about the value of road verges and power-line corridors for landscape-scale diversity and connectivity. This will be streamed live on zoom at 13:15 CET on Tuesday 28th February 2023.
Juliana is interested in the relative effects that different land use practices (e.g. agricultural land, productive forests, semi-natural grasslands) have on biodiversity and on how these effects change at different spatial scales. Also, Juliana has an interest in developing strategies for biodiversity conservation that account not only for local disturbances but also for human-induced changes in the landscapes.
Lutz Eckstein is the lead author of a paper summarizing the current knowledge on the biology of the invasive legume Lupinus polyphyllus Lindley. The paper has recently been published in the journal Perspectives in Plant Ecology, Evolution and Systematics in the series “Biological Flora of Central Europe” (https://doi.org/10.1016/j.ppees.2022.125715). This work is a cooperation with Erik Welk (Martin-Luther-University Halle-Wittenberg and German Centre for Integrative Biodiversity Research (iDiv) Halle, Germany), Yves Klinger and Wiebke Hansen (both Justus Liebig University Giessen, Germany), Tommy Lennartsson and Jörgen Wissman (both Swedish University of Agricultural Sciences (SLU), Uppsala, Sweden), Kristin Ludewig (University of Hamburg, Germany), and Satu Ramula (University of Turku, Finland).
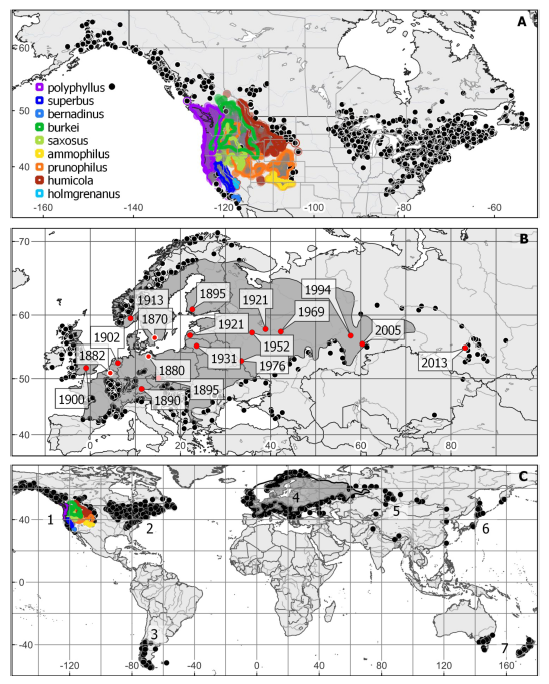
The paper gives a thorough review of the species’ taxonomy, presents distribution maps for North America, Europa and the world (Fig. 1), illustrates the life cycle of L. polyphyllus, and it contains a comprehensive discussion of potential management options. During the research for this review, the authors encountered some doubtful information about L. polyphyllus that uncritically reiterates in several fact sheets, reports and webpages. One such erroneous piece of information refers to the apparently very high longevity of seeds, which was taken from a modelling study on seed longevity under optimal dry and cold storage conditions. Similarly, there is some uncertainty and large variation concerning the actual lifespan of the species. Another piece of doubtful information is the deep rooting depth of L. polyphyllus, which may rather characterize a maximum than a representative average value. Finally, the species is sometimes considered a “rhizomatous perennial” although it lacks true rhizomes. These points highlight some critical knowledge gaps, which partly relate to aspects of the species’ life cycle and morphology that may be either time-consuming or labor-intensive to study.
Fig 1. Distribution of Lupinus polyphyllus s.l. (A) In North America, the native segregates in the west partly overlap in their distribution and are delimited by outlines according to the color scheme in the legend. Non-native, synanthropic occurrences are indicated by black dots. Distribution data based on digitally available herbarium specimen locations and county records (for data sources see Table 1 in the paper). (B) In Europe, numbers give the first records for the species in different countries/regions (cf. Table 6 in the paper). (C) Numbers refer to the textual descriptions (for details, see the paper) of the non-native naturalized distribution across the globe
The authors conclude that there is currently no evidence-based strategy for a cost-efficient management of L. polyphyllus. The development of such control measures is necessary because L. polyphyllus is among the most problematic non-native plant species in Europe with respect to environmental and socio-economic impacts. The species has significant negative effects on community structure, composition, species richness and diversity, especially in nutrient-poor habitats such as alpic mountain hay meadows, alpic mat-grass swards but also nutrient-poor road verges or riparian terraces.
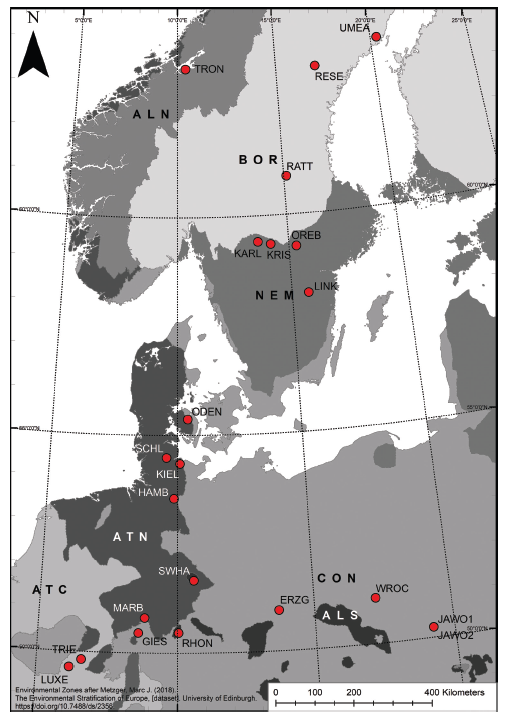
Lutz Eckstein is the senior author of a multi-author paper on the phenology and morphology of the invasive legume Lupinus polyphyllus along a latitudinal gradient in Europe. He has also been the project coordinator, while Kristin Ludewig (Hamburg University) has led the preparation of the paper, recently published in the journal NeoBiota (https://doi.org/10.3897/neobiota.78.89673). In total, 30 authors (researchers, students, consultants, conservation agency employees) have been involved in this project and the publication. Each co-author contributed data of a lupine population and the whole project covered a >2000 km long latitudinal gradient including the countries Luxembourg, Germany, Poland, Denmark, Sweden and Norway (Fig. 1).
Fig. 1. Position of the 22 study sites (for site abbreviations, see Table 1 in the paper). Background map depicts climatic zones. ATC – atlantic central, ATN – atlantic north; ALS – alpine south; CON – continental; NEM – nemoral; BOR – boreal; ALN – alpine north. For certain analyses sites in BOR and ALN were pooled.
Plant phenology is the timing of seasonal events, such as budburst, greening, flowering, and fruit ripening. Phenology influences the fitness of individual plants, controls species distribution ranges, and may have cascading effects on communities and ecosystems. Temperature is one of the most important drivers of plant phenology together with day length. However, the potential for adaptation of phenology may also be key to understanding the success of invasive plant species, which will benefit from ongoing global change. Lupinus polyphyllus Lindl. (Fabaceae) is a perennial herbaceous hemicryptophyte originating from western North America that was introduced in Central Europe as an ornamental plant in the 19th century. From Central Europe and Scandinavia, the species spread very successfully to almost all parts of Europe, now ranging from the Pyrenees in the West to the Ural (and beyond) in the East (Eckstein et al., unpublished data). From North to South, L. polyphyllus is currently covering the full range of Europe, except for Mediterranean zones such as the Iberian Peninsula and Italy.
The overall aim of the paper was to understand how the timing, temperature dependence of flowering and fruiting, and performance (canopy height, potential seed production and seed release height) of L. polyphyllus change along the latitudinal gradient from Central to Northern Europe. The authors tested differences between populations from different climatic zones and quantified variation in phenology in relation to latitude. The present study is probably the first attempt to quantify variation in phenology of an invasive plant across a large latitudinal gradient in the field.
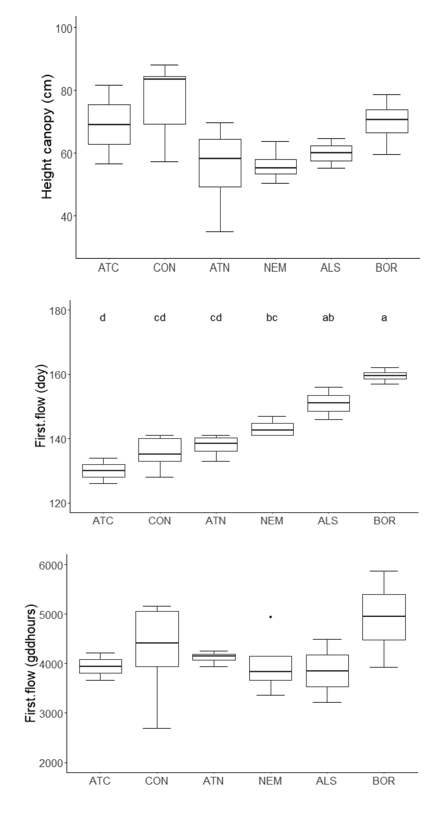
The paper studied variation in growth and phenology of flowering and fruiting of L. polyphyllus using measurements and >1600 digital photos of inflorescences from 220 individual plants observed weekly at 22 sites. While canopy height of plants did not vary significantly along the latitudinal gradient, the day of the year (doy) at which different phenological phases were reached, increased 1.3–1.8 days per degree latitude (Fig. 2), whereas the growing degree days (gdd) required for these phenological phases decreased 5–16 gdd per degree latitude. However, this difference disappeared, when the day length of each day included in the calculation of gdd was accounted for (Fig. 2). The day of the year of the earliest and the latest climatic zone to reach any of the four studied phenological phases differed by 23–30 days and temperature requirements to reach these stages differed between 62 and 236 gdd. Probably, the invasion of this species will further increase in the northern part of Europe over the next decades due to climate warming. For invasive species control, the results suggest that in countries with a large latitudinal extent, the mowing date should shift by ca. one week per 500 km at sites with similar elevations to target the species in the same phenological phase.
Fig. 2. Summary figure showing canopy height (cm), day of the year (doy) of first open flowers and growing degree hours (gddhours) first open flowers of populations of L. polyphyllus in different climatic zones. For abbreviations of climatic zones, see Fig. 1.
RivEM – Karlstad University
… is a research group associated to the subject of Biology and the Department of Environmental and Life Sciences at Karlstad University, Sweden. We conduct both basic and applied research on human impact on river ecosystems, and how this impact can be minimized.
In Swedish the research group is called Naturresurs Rinnande vatten (Acronym NRRV, hence the url nrrv.se).