Jacqueline Hoppenreijs, one of the RivEM PhD students, will be giving a seminar entitled Effects of hydrogeomorphology on riparian seedbank and vegetation composition. Join us on 13 June (13:15 CEST) on Zoom via https://kau-se.zoom.us/my/magnuslovenwallerius or in room 5D306 on KAU Campus.
This post covers chapters 16 & 17 from Futuyma & Kirkpatrick’s book on Evolution (2018). The author of this post is Sam Shry.
Phylogeny: how we are all connected
We know that we look different from a crocodile (well, most of us), but how did we diverge from our common ancestors millions of years ago? Can we trace back our common ancestry and how we are related to every other organism on this planet via our genetic relationship and morphological characteristics? That’s what phylogeny tries to understand and with the help of modern genetic analysis, this process has opened our eyes to understanding the phylogenies of every species.
By backtracking each species’ ancestry, we can understand how they are all related and can determine when species split based on morphology, derived features, and genetic architecture. Understanding the exact order of evolution among species can be tricky, as mutations can occur repeatedly, creating sometimes the same evolved characteristics multiple times in multiple phylogenies (homoplasy). One example is the evolution of winged insects. It is a convoluted mess of where and when each species developed or lost wings, making the phylogenetic tree hard to disentangle when basing the entire analysis only on the presence or absence of wings. To avoid these issues, it is important to analyze more than one characteristic when determining relationships and by combining different tools for analysis (morphological differences, genetic differences, etc.). It also depends on the temporal scale; recent evolutionary differences need to be analyzed using faster-evolving parts of the genome, whereas longer time scales require slower-evolving genome sections. What is also important to remember is phylogeny and genealogy of species may not match up completely as groups of genes can be copied and inherited even with speciation, called incomplete lineage sorting. There can also be problems like introgression, where regions of one genome are taken from another species via hybridization, horizontal gene transfer, etc., which can give a misleading picture of the species’ genome. The best way to avoid these problems with differentiating species is to analyze larger regions of the genome to detect more variability between species.
DNA sequencing is done in multiple ways, but one of the most common ways is parsimony, examining how different species are related to one another based on their genetic base changes, simply put, the phylogenetic tree requiring the fewest evolutionary changes. It can, however, do a bad job of parsing the exact phylogeny of where and when individuals split into new species. More robust, statistical methods such as likelihood estimations and Bayesian inference do a better job of differentiating species exactly where and when points of speciation occur. Though, DNA sequencing is difficult when DNA is hard to come by, as with extinct species. This is when old-school methods are still used to construct phylogenies from morphological data instead. In both methods, we are trying to pinpoint the exact time in history when evolutionary changes occurred, so understanding the “when” is an important aspect. We can do this by analyzing the molecular clock within DNA sequences as we have learned about previously, giving us a rate of sequence evolution, which can be used to estimate the time of divergence. Understanding when and where evolutionary changes occurred gives us a road map to exploring reasons for divergence.
We can use phylogenies to answer questions about mutations, adaptations, and genetic variation in general. One example is the three-spined stickleback, which has invaded freshwater systems of almost all reaches of the Northern Pacific and Atlantic oceans. Their variation in armored plates along their back is quite diverse and has been traced back to a single locus, creating this adaptation throughout freshwater populations. Phylogenies can also be used to trace almost any species’ characteristics, such as language, tails, and even virus prevalence. Phylogenies can be compared to one another, especially when investigating adaptions. Investigating the adaptive coupling of characteristics while controlling for phylogenetic relations is a valuable tool for understanding the evolutionary relationship between species’ characteristics.
Figure 1. Three-spined stickleback, Gasterosteus aculeatus. Photo by Jack Wolf
Of course, one of the main purposes of developing phylogenies is to classify species into groups and taxa, as has been done from Aristotle, to Linnaeus to Darwin, to modern phylogenetic classifications. Conveying the relationship between species is an important step to understanding biodiversity and leads to further exploration of genetic architecture passed down within and between species.
The history of life… really, that’s the title
A bold title and difficult to summarize into two pages, the underlying connection between today and earth hundreds of millions of years ago are the species that have inhabited this planet over time. We can follow a species through time using their fossils as stepping stones in their evolution, while simultaneously trying to understand how the earth’s environment has changed and impacted species’ biodiversity. As the earth has changed, for example via plate tectonics, the layering or strata of sediment holds records of past organisms and gives us the ability to set a time scale to when organisms were found and when they were lost. These fossils are highly valuable, but very difficult to come by, making our fossil record patchy and incomplete. After the “big bang” 14 billion years ago (Gya), biotic “Life” was formed by abiotic chemical reactions (around 3.5 Gya), creating simple organic molecules. The formation of molecules that could replicate independently (most likely RNA) allowed for evolution by natural selection to begin to shape variation. The complexity of life continued to expand and develop over billions of years, from asexual unicellular to sexual unicellular to multicellular organisms. The Precambrian era (Table 1) brought about evolved photosynthesis and oxygen to the atmosphere, as before this period all life (prokaryotes from the groups Archaea and Bacteria) were anaerobic. The emergence of eukaryotes also occurred (1.8 Gya), opening greater possibilities for diversity.
Table 1. Each step in organic life history and a few important evolutionary points from the millions of years in between.
Time period
Important stuff that happened
Precambrian: 2.5 Gya – 541 Mya
-photosynthesis evolved -oxygen to atmosphere -emergence of eukaryotes
Cambrian: 541 Mya – 485 Mya
-explosion of diversity -development of the genetic toolkit for animals -vertebrates arise -ended with large extinction event
Paleozoic: 541 Mya – 252 Mya
-species diversity -evidence of modern phyla -movement to land and terrestrial animals -plant diversity and terrestrial modifications -early ancestors of mammals -ended with mass extinction
Mesozoic: 252 Mya – 66 Mya
-age of reptiles -birds -break up of Pangea -slow recovery of diversity – adaptive radiation -angiosperms and insect diversity -End with mass extinction via asteroid impact
Cenozoic: 66 Mya – today
-modern time -closely related ancestors to modern day organisms -recovery and increased diversity -forming of modern continental geography and climate -adaptive radiation of mammals -human speciation
The Cambrian era of only about 55 Mya, but created large-scale animal diversity by introducing many new species and classes of animals. This explosion of diversity is attributed to new feeding abilities and novel ways of living. Ecological changes, such as increased atmospheric oxygen and a warmer climate could have also helped in the rise of diversity. More diversity during the Paleozoic period meant an increase in aquatic organism diversity, creating larger and more complex invertebrates and vertebrates. Large squids, sea stars, and molluscs were conceived and some of the first reefs were built by two coral groups. The large extinction event during the late Cambrian gave rise to large-scale diversification with new phyla with new ways of life. Boney fish also arose during this time, creating the ancestors of today’s fish species. Aquatic plant diversity also exploded and advanced to form terrestrial plants with roots that form in the newly acquired terrestrial biomass of organic soil. Insects soon followed, as herbivores feeding on terrestrial plants. The transition from fish to tetrapod to terrestrial vertebrates also occurred during this time, moving from fin to limb. At the end of this era however, a mass extinction event occurred as land masses redistributed themselves, creating chaotic and unfavorable living conditions for many species. The Mesozoic era brought about more stable conditions for living and allowed for yet another round of adaptive radiation and diversification with the rise of new reptile species, both marine and terrestrial. Insects and plants diversified, with angiosperms bringing about insect diversity. There was an explosion in terrestrial insect diversity and the sheer size of some is mindboggling, for example, the Arhropleura, a millipede that reached 2.3m long. Birds also came to be with the diversity in reptiles leading to their wing adaptations for flight. The end of the period is marked by another mass extinction due to the impact of an asteroid off the coast of the Yucatan Peninsula, blocking the sun and creating a long-lasting winter. After that, the Cenozoic era begins around 66 Mya and marks the modern time in evolutionary history. The period is marked by the forming of modern-day continents, climate, and species diversity. The re-diversification formed what are today’s species and the radiation of mammals, eventually leading to the speciation of humans. During this period we also see the first indications of human impact on species diversity, with the extinction of what were called megafauna or large-bodied mammals such as mammoths.
As we can tell from this chapter, life has been ever shaped by the climate and environmental changes on Earth, era after era. Extinctions give rise to adaptive radiations with new species adapting to the present environmental conditions and ever-advancing in complexity. Though today, many of the extinctions occurring are due to human impacts. From our microscopic time span of existence, we are causing rapid global change to both environments and biodiversity. We will just have to see how evolution and Earth sort us out over the next million years…
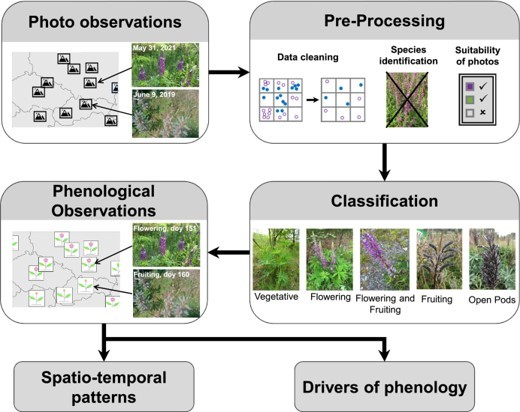
Lutz Eckstein is co-author of a paper presenting a workflow for iPhenology (Fig. 1), i.e., the use of publicly available photo observations to track phenological events at large scales. The paper, which has recently been published open access in the journal Methods in Ecology and Evolution (https://doi.org/10.1111/2041-210X.14114), is led by Yves Klinger (Justus-von-Liebig University Gießen, Germany), who will be visiting Biology at KaU in June 2023. Overall, publicly available citizen science photo observations are suitable to track key phenological events and can thus significantly advance the knowledge on the timing and drivers of plant phenology. In future, integrating the workflow with automated image processing and analysis may enable real-time tracking of plant phenology. To learn more about what sparked the idea for this paper and what may be advantages of working with citizen science photos, see the Methods in Ecology and Evolution blog (https://methodsblog.com/2023/05/22/using-citizen-science-photos-to-perform-phenological-studies/).
Figure 1. Proposed workflow for iPhenology. First, observations are pre-processed by removing problematic observation and, if necessary, reducing spatial aggregation. Second, photos are checked for correct identification and suitability before being classified. Unsuitable or misidentified photos are removed. For the resulting phenological observations, there are many potential uses.
This post covers chapters 14 & 15 from Futuyma and Kirkpatrick’s book on Evolution (2018). The author of this post is Jeff Marker.
The Evolution of Genes and Genomes
One of the most common sources of evolutionary adaptation is the emergence of new genes. In eukaryotes, the most common mechanism for the emergence of new genes is by gene duplication, which occurs when there is an error in DNA replication. Gene duplication can result in the creation of a new gene copy, which can then evolve to acquire new functions or to specialize in one of the functions of the original gene. The result is a gene family, which is a set of loci that originated by gene duplication and that typically have related biochemical roles. Gene families can lead to the emergence of new biochemical pathways and new structures. For example, in humans, the amylase gene family has been duplicated and evolved to produce different amylase enzymes, allowing humans to digest a wide range of starches from different plants (Perry et al., 2007).
Whole genome duplication is much rarer than gene duplication, but is a key mechanism for the evolution of the genome in many groups of organisms, especially plants. Whole genome duplication can result in the doubling of the number of chromosomes in an organism, which can then allow for the emergence of new gene functions through the duplication and divergence of gene families. For example, the multiplication of complete sets of chromosomes that have occurred in the Brassicaceae family has resulted in the emergence of many new gene functions and the evolution of novel morphological traits (Bancroft et al., 2011).
Other mechanisms of gene emergence include exon shuffling and appearance of de novo genes. Exon shuffling is a process where coding regions (exons) from different genes are rearranged and recombined, creating new genes with unique functions. De novo genes originate from non-coding regions of the genome, evolving independently of existing genes. They contribute to biological functions and evolutionary innovation through mechanisms like mutation, selection, and gene duplication.
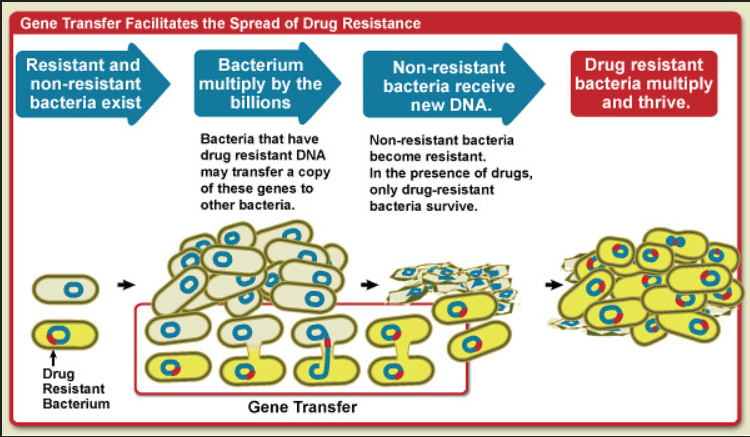
Another mechanism for the emergence of new genes is horizontal gene transfer (HGT), which involves the possession of genes from unrelated species (Figure 1). HGT is particularly common in prokaryotes, where it has enabled the rapid evolution of traits. For example, the spread of antibiotic-resistance genes among bacteria has been assisted by the HGT of plasmids carrying resistance genes (Davies and Davies, 2010).
Figure 1. Diagram showing how gene transfer facilitates the spread of drug resistance. Credit: National Institute of Allergy and Infectious Diseases
In contrast to the previously mentioned methods of genome evolution that saw the creation of genes, chromosomal deletions can eliminate functioning genes contributing to both “good” and “bad” genomic outcomes. Natural selection can cause a deletion to increase in frequency if the deleted gene codes for a protein that increases the risk of infection. For example, a deletion in the CCR5 gene that codes for a cell surface protein used by the HIV virus to enter human cells has become fixed in some human populations.
The continuing evolution of genomes leads to a situation where some species end up with a very high number of genes. For example, the human genome consists of approximately 3 billion base pairs. However, only a small fraction of our genome is used to encode proteins and regulate gene expression. Estimates suggest that we only use about 2% of our genome to code for proteins or other gene products. The rest, termed “junk DNA,” includes regions with regulatory elements and non-functional remnants known as pseudogenes. Pseudogenes are genes that have lost their protein-coding ability through mutations. Their presence aligns with the neutral theory of molecular evolution proposed by Motoo Kimura. According to the neutral theory, the majority of genetic changes are caused by random genetic drift rather than selective pressure, resulting in non-functional sequences like pseudogenes. Understanding the significance and potential functions of these non-coding regions remains an active area of genomic research.
Evolution drives changes in gene expression patterns that can affect the survival of organisms ultimately leading to new traits and new species. Changes in gene expression occur through alterations in transcription factor binding sites, alternative splicing patterns, and epigenetic changes to DNA and histones. One example of the evolution of gene expression in insects is the wing pattern of Heliconius butterflies. Their diverse wing colors aid in reproduction and to signal to predators that they are unpalatable. Researchers found that changes in gene expression in the developmental pathway that produces the wing patterns of Heliconius butterflies have contributed to their diverse color patterns (Figure 2). The study showed that the expression of a transcription factor gene called optix, which is involved in forming the eyespot pattern on the wings, has evolved differently in different species of Heliconius butterflies (Martin et al., 2012).
Figure 2. Some of the different patterns of Heliconius butterflies. Credit: A. Meyer
Coding regions, gene expression, chromosome structure, and genome size are interconnected processes shaping an organism’s complex life. Changes in coding regions can lead to the emergence of new functional genes or alterations in protein structure. Concurrently, adjustments in gene expression patterns can drive phenotypic variation. Chromosome evolution plays a role in these processes, as rearrangements and other changes reshape the distribution of genes within a genome. Furthermore, changes in genome size can influence gene regulation, complexity, and the evolutionary trajectory of a species.
Evolution and Development
Evolutionary developmental biology (EDB) aims to integrate information from embryology, developmental genetics, and population genetics to understand how genes and changes in genes are expressed as phenotypes and changes in phenotypes. By understanding the mechanisms that produce phenotypes, we can better grasp how they evolve. Mutational changes in the genes that produce a developmental pathway may cause advantageous alterations in the phenotype, leading to the evolution of both the phenotype and its underlying genetic network.
Differences among species often result from changes in the relative developmental rates of different body parts or in the rates or durations of different life history stages, which are known as allometry and heterochrony. Some characteristics have evolved by heterotopy, where the expression occurs at a novel location on the body. For example, the inner ear bones of mammals were originally found as part of the jaw in distant common ancestors. These bones co-opted to function in the ear away from the jaw and helped mammals become more effective at hearing, aiding their survival and reproduction.
The vast diversity of multicellular eukaryotes is due to the diverse uses of a toolkit of genes and developmental pathways. Developmental pathways include signaling proteins, enhancers, transcription factors, and structural genes. Evolutionary changes in the regulatory connections among signaling pathways and transcription factors are believed to underlie much of the phenotypic diversity seen in nature. These regulatory networks coordinate the interactions among genes and other molecules to control gene expression. Regulatory networks govern biological processes and organism responses to environmental cues. One example of a gene regulatory network is that involved in the development of fruit flies. This network includes genes such as the Hox genes, which control the body segment identity, or where the different body segments are supposed to line up. The Hox genes regulate each other’s expression through a series of complex interactions, forming a network that ensures the proper patterning and differentiation of different body segments during development.
During evolution, genes and developmental pathways are often co-opted for new functions, a process that is probably responsible for the evolution of many new traits. This process results from evolutionary changes in functional connections between transcription factors and cis-regulatory elements. Modularity among body parts is achieved by patterning mechanisms whose regulation is often specific to certain structures, segments, and life history stages. Modularity helps different parts of the body develop divergent morphologies. Pleiotropic effects of genes that affect functionally interacting characteristics may evolve, resulting in the evolution of functional modules known as phenotypic integration. One example of phenotypic integration is the correlation between beak size and body size in birds. In many bird species, larger birds tend to have larger beaks, and smaller birds tend to have smaller beaks. This is an example of phenotypic integration because the size of the beak and the size of the body are functionally related traits that are influenced by overlapping sets of genes.
Genetic and developmental constraints can make some imaginable evolutionary changes unlikely. Based on changes in the expression of certain genes and developmental pathways in response to environmental signals, a single genotype may be expressed as an array of different phenotypes, known as the genotype’s norm of reaction. Reaction norms are genetically variable and can evolve by natural selection. If an environment varies, phenotypic plasticity may evolve. Conversely, selection for a constant phenotype can result in canalization. Genetic assimilation is the genetic fixation of one of the states of a phenotypically plastic character. It is not known how important genetic assimilation is in evolution, nor is it known if adaptation may occur first by a non-genetic phenotypic change that later becomes genetically fixed by natural selection.
The evolution of genes and genomes involves complex processes such as gene duplication, whole genome duplication, exon shuffling, de novo gene emergence, and horizontal gene transfer. These mechanisms contribute to the emergence of new functions and the huge amounts of genetic diversity we see in the world. Changes in gene expression patterns play a vital role in evolutionary adaptation and the development of new traits. Chromosome evolution and alterations in genome size further shape the evolutionary course of species. Understanding that these processes are firmly connected provides insights into the complexity and diversity of life forms that have evolved on our planet.
References
Bancroft, I., Morgan, C., Fraser, F., Higgins, J., Wells, R., Clissold, L., … & Trick, M. (2011). Dissecting the genome of the polyploid crop oilseed rape by transcriptome sequencing. Nature biotechnology, 29(8), 762-766.
Davies, J., & Davies, D. (2010). Origins and evolution of antibiotic resistance. Microbiology and molecular biology reviews, 74(3), 417-433.
Martin, A., Papa, R., Nadeau, N. J., Hill, R. I., Counterman, B. A., Halder, G., … & Reed, R. D. (2012). Diversification of complex butterfly wing patterns by repeated regulatory evolution of a Wnt ligand. Proceedings of the National Academy of Sciences, 109(31), 12632-12637.
Perry, G. H., Dominy, N. J., Claw, K. G., Lee, A. S., Fiegler, H., Redon, R., … & Stone, A. C. (2007). Diet and the evolution of human amylase gene copy number variation. Nature genetics, 39(10), 1256-1260.
Peter Lambert (A Marie Curie Postdoctoral fellow at the Environmental and Life Sciences Department of Karlstad University) will be giving a seminar entitled Actions for Insects – from insect decline to actions for insect conservation. You can join live on zoom via https://kau-se.zoom.us/my/magnuslovenwallerius or in room 5D307 on KAU Campus.
This post covers chapters 12 & 13 from Futuyma and Kirkpatrick’s book on Evolution (2018). The author of this post is Jacqueline Hoppenreijs.
In blog 4, we briefly touched upon one of the advantages of dispersal: it decreases competition for resources with your nearest and dearest. Residency can have multiple outcomes that vary from extremely cooperative, via neutral, to extremely violent.
Cooperation and conflict
When two individuals coexist, one can help the other (“cooperate”) through altruism or mutualism, or do the opposite (“conflict”). Altruism is probably the most renowned example of these interactions, and indicates that one individual sacrifices something to another’s benefit. In the case of mutualism, there are only winners: everyone benefits. Conflict can consist of direct harm, but also indirect actions such as cheating. That’s not to say that a plant and a bee are sitting down for a game of Monopoly, but it might look like pollinators that are not doing their job by bypassing a plant’s anthers and stealing the nectar (Figure 1).
Figure 1. Pollinators robbing plants of their nectar instead of taking the front door and take some pollen with them.
Now, it is tempting to think that those that cheat will win from the others, because they get more resources with less effort, and might thus be able to withstand selection pressure better. But wouldn’t populations come to consist exclusively of cheaters and then collapse? Not quite, so there must be another mechanism at work. That mechanism is not group selection, which was a popular hypothesis until the 1960s and assumed that selection pressure didn’t work on the individual but on the level above. There is, however, a bunch of other mechanisms through which evolution has resulted in cooperation rather than conflict.
A well-known example of non-family cooperation is to find safety or success in numbers. This behaviour can directly increase the chance of not-being-eaten, or increase the chance of catching a prey. Other types of cooperation take more time to pay off, or only work as such because the individuals involved interact repeatedly. The latter means that actions are reciprocal, as opposed to one-off selfish behaviour. This might look like individuals copying each other’s behaviour or copying their own, previous behaviour until that’s not rewarding anymore. A series of cooperative interactions can also change if a helping individual decides to punish a cheater for being selfish. Or even worse, when one individual damages another at a cost for itself! All of this changes when family enters the game. Now, there’s not just profit for yourself and increased direct fitness to be gained, but you can gain by helping others survive. Why? Because they’re likely to be carrying the genes you carry, which means there’s an indirect fitness advantage to be gained! Kin selection like this is advantageous if the relatedness and benefit for the recipient outweigh the costs of the helping individual.
Conflict within family, on the other hand, can take some really extreme forms. That starts at the parental level: we’ve heard about male-male competition in blog five, but males can go as far as to harm the female they’re mating with (called sexually antagonistic competition) in order to increase their own reproductive success. The female praying mantis’ sexual cannibalism barely makes up for all the suffering her sisters go through. A less deadly form of parental conflict is the distribution of parental care to the offspring, which takes time and energy from either or both parents.
Moving onto the next generation, we see that there are parents that kill other individual’s offspring or even their own, which is called infanticide. The former makes sense in that they are making space for their own genes in the population by removing another individual’s genes, and the latter might make sense if there’s resource limitations. The same reasoning might apply when siblings compete for the same resource, which is called siblicide. This touches upon the conflict that exists between parents and their offspring. While one or few offspring are usually selfish as they profit most when they themselves survive, it’s in the interest of the parent to produce as many successful offspring as possible.
After so much dysfunctional family dynamics, we’re moving to animals that have taken family relationships to a whole other level: eusocial animals, such as many bees and termites. Here, we see the positive side of kin selection, in that infertile offspring all increase their indirect fitness by taking care of their mother, the nest, or helping rear their (half) siblings. There’s a, for lack of better words, negative side, too. To keep the relatedness with reproductively successful offspring high, eusocial animals don’t hesitate to disadvantage some family members that are not as related to them as others.
We’ve seen that conflict can exist between individuals because they want to increase their direct or indirect fitness. It also exists within individuals, for example when alleles kill off other alleles during meiosis, leading to segregation distortion, or in the form of transposons, bits of DNA that keep replicating themselves within a genome. Conflicts between mitochondrial and nuclear DNA, which are spread via the maternal line and both parents respectively, also occur widely. It’s in the mitochondrion’s interest to increase female production also when it comes at a cost to male production, so the nuclear DNA has to jump into an arms race to try and undo any changes that might disadvantage male reproduction. The previously mentioned concept of group selection shouldn’t get dismissed entirely, as there are examples of traits evolving in groups that are not necessarily advantageous for its individuals. Groups of individuals with such benevolent traits survive better than groups without them. Benevolence doesn’t always mean pure altruism, by the way: lower production of toxins by plants (Wilson, 1987) and “less-deadliness” of pathogens are well-described examples, too. Especially pathogens want to be careful: they’ll want to spread as much as they can, but there’s little use in killing off the entire host population since that means they’re doomed themselves. Units that depend on vertical transmission, that is reproduction as opposed to horizontal transmission/infection, profit from making their host as successful as possible. Not so surprising then, that they are the driving forces behind some of the major transitions that our planet has seen. The adoption of mitochondria and cyanobacteria have led to successful single-cell organisms. Some of these have, through mechanisms similar to kin and group selection, been able to transform to what we know as multicellular organisms today.
Interactions among species
Neither single-cell nor multicellular organisms exist in a vacuum or only interact with organisms of their own kind. Species interactions are variable over time, in how much species depend on each other and in its effect on the respective participants of the interaction. Examples include a specific insect and plant needing each other a few weeks per year for food and pollination respectively, to a parasite that spends its entire life in its host and depends on it for all its food and the dispersal of its offspring. Close associations are called symbiosis and species that are very attuned to each other, such as corals and zooxanthellae (Stanley & Swart, 1995), are said to be co-evolved. This means that they have undergone reciprocal genetic change because they’ve exercised selection on each other.
Not all of these interactions are positive; becoming the host to a parasite or a prey to a predator usually doesn’t bode well for an individual or a population. Both the enemy and the victim in these relationships need to evolve fast enough and in such a way that they can outperform the other; this can turn into an evolutionary arms race if both are under selection for the same trait. Co-evolution doesn’t have to be unidirectional, as enemy and victim can follow each other to increase and decrease trait values over time.
There are multiple strategies for escaping or outperforming enemies and becoming successful in this arms race. Many are visual-based: by displaying warning signals of being dangerous (aposematism) or mimicking a dangerous species (mimicry). If you’re not a dangerous species yourself (Batesian mimicry), you disadvantage the actually dangerous species by teaching enemies that risks are low. Being dangerous yourself and mimicking a dangerous colleague (Müllerian mimicry), on the other hand, reinforces the defence mechanism. Other strategies can be olfactory or taste-based, such as in plants that produce chemical compounds to protect themselves from herbivory. Throughout history, such successful adjustments have often been seen to lead to quick radiation and many new species with that adjustment, both on the enemy and the victim side. Whether a parasite can successfully infect its victim, can depend on whether the victim can recognise and resist (gene-for-gene model), or whether the parasite matches the victim’s genetic profile (matching allele model). Parasites and pathogens have been shown to adapt relatively quickly to their victims, which means that the above-mentioned shift towards benevolence doesn’t always take place. This depends, amongst others, on the genetic diversity on both the enemy- and victim-side and the way in which the parasite or pathogen is transmitted and received. The latter can be as simple as the difference between vertical and horizontal transmission, but can also have to do with the life cycle of host, pathogen and potential intermediate hosts. In the case of malaria, for example, humans and mosquitoes are hosts of most Plasmodium species, which is complicated as a life cycle in itself. There seems to be at least one Plasmodium species, however, that has found another intermediate host in macaque monkeys, complicating the transmission process further (Centers for Disease Control and Prevention, 2020).
Moving on to the sunnier side of life, there are also a lot of species interactions from which one or both species profit. This mutual exploitation, can, just as with the cheating family or population members that we saw earlier, go awry when one of the participants starts cheating the other. That can lead to immediate sanctions being taken (e.g. lower rewards for the cheater) but can also continue to exist because cheating happens relatively little or there are non-cheaters to compensate potential losses. Even here, one could speak of an arms race where both participants exploit the other as much as they can, and increasingly over time.
Not all species interactions are one-on-one, or as close as some of the previously-mentioned examples. Very often, species interact simply because they use the same space or other resources. If there are limited resources, however, this may lead to competition. One consequence of competition might be that one or more species is driven to extinction. This principle, and the related “Diversity Paradox”, have been the topic of many studies (Simha et al., 2022). A second consequence is the selection-by-lack-of-resources that can cause divergence of resource use and may lead to the evolution of new species. If species that overlap in their use of resources co-occur, they might each display more or different resource-use compared to when they occur alone. This is called ecological release, but shifts to interference competition when one of the species starts to affect its competitor to keep it from using the resource in question. If this is done by a species not native to the area, one can speak of invasive alien species (Figure 2).
Figure 2. Competition for light and space in a Swedish riparian zone by the American Skunk-cabbage (Lysichiton americanus). Photo by Owe Nilsson.
Whether friends or foes, newcomers and the species that already were in a certain place together form an ecological community. The local environment and its inhabitants impose a “filter” on the species that occur in a larger area to decide which of them manage to live in a certain spot. The mechanisms of competition can lead to evolutionary divergence, whereas the environmental filtering can lead to evolutionary convergence. That’s why we often see functionally similar, but taxonomically different species communities in ecologically similar, but geographically different areas. Uniting community ecology and evolutionary biology helps us discern these patterns and the eco-evolutionary past and present.
References
Centers for Disease Control and Prevention. (2020). Malaria. (DPDx – Laboratory Identification of Parasites of Public Health Concern). https://www.cdc.gov/dpdx/malaria/index.html
Futuyma, D., & Kirkpatrick, M. (2018). Evolution (4th ed.). Oxford University Press.
Simha, A., Pardo-De la Hoz, C. J., & Carley, L. N. (2022). Moving beyond the “Diversity Paradox”: The Limitations of Competition-Based Frameworks in Understanding Species Diversity. American Naturalist, 200(1), 89–100. https://doi.org/10.1086/720002
Stanley, G. D., & Swart, P. K. (1995). Evolution of the Coral-Zooxanthenllae Symbiosis During the Triassic: A Geochemical Approach. In Paleobiology (Vol. 21, Issue 2, pp. 179–199).
Wilson, J. B. (1987). Group selection in plant populations. Theoretical and Applied Genetics, 74(4), 493–502. https://doi.org/10.1007/BF00289828
This is a literature course on the book Evolution by Douglas J. Futuyma & Mark Kirkpatrick (Fourth Edition) during the first half of 2023. This write-up, authored by Louis Addo covers chapters 10 to 11.
All about Sex
Anisogamy, often known as sexual reproduction which involves large immobile eggs and small mobile sperm are present in plants, animals, and certain eukaryotes. Ansiogamy also describes the dimorphism in the sizes of gametes which defines the different sexes. When suitable partners are few, hermaphroditism is advantageous since the different sexes employ different methods to determine the sex of the embryos. Sexually dimorphic characteristics may not evolve due to genetic limitations, but they may nevertheless be produced by sexually antagonistic selection. Certain species, including deep-ocean anglerfish, have males that adhere to females and transform their bodies into practically complete testes to aid in fertilization.
Charles Darwin’s theory of sexual selection states that competition for mates among members of one sex results in the evolution of secondary sexual traits and mating habits. According to Bateman’s theory, the number of females a male mates with determines how successful his reproduction is, amplifying males’ secondary sexual traits in the long term. There are two major modes of sexual selection: selection by male-male competition and selection by female choice. In male male-male selection, males directly compete through interference or resource control. Males may acquire characteristics and behaviors such as horns and battles to get access to females. Another type of sexual selection is female choice, in which females can “choose” partners based on particular traits. For example, female salmon will choose the largest and strongest males among the options. In other mating systems called sex role reversal, females can actively seek males with males exhibiting parental behavior such as taking responsibility for incubation and rearing of the chicks. This is shown in Figure 1 where Two female red phalaropes (Phalaropus fulicarius) fight over the smaller, duller-plumaged male on their breeding ground. In contrast to most birds, female phalaropes court males, which care for the eggs and young in their nests.
Figure 1 Sex role reversal shown by Two female red phalaropes (Phalaropus fulicarius) fight over the smaller, duller-plumaged male on their breeding ground (Note from Futuyma, D. J., & Kirkpatrick, M. (2017). Evolutionary. Evolution (Fourth ed.). pp. 254. Sunderland, Massachusetts: Sinauer Associates, Inc.)
Sexual selection is influenced by variables including resource availability and operational sex ratio. Flowering plants also engage in sexual selection, with the rivalry between pollen grains and beautiful flower displays. Direct selection on choice occurs when genes that affect preference also directly affect survival or fertility. Indirect selection, on the other hand, is when preference genes are connected to other genes that are spreading due to selection. Understanding the processes and results of sexual selection can provide insight into how secondary sexual traits and mating habits have evolved across a range of organisms.
Sex ratios, or the ratio of males to females, are typically equal at birth in most creatures with distinct sexes because meiosis in males typically transmits the X and Y chromosomes with equal likelihood. In some species, sex is determined not by chromosomes but by physical or social environment. This is known as environmental sex determination. In the insect order Hymenoptera, which includes ants, bees, and wasps, a female either lay an unfertilized egg that develops into a male or fertilizes an egg that develops into a female to decide their sex. There are several exceptions to the rule that natural selection favors equal sex ratios, such as fig wasps, where considerably more females than males are born. As fig wasps are often a single family of brothers and sisters, this tendency may be explained by considering how organisms within them are linked. A mutation that enables females to bear male offspring will be selected because only females leave the fig to start the next generation.
In asexual reproduction, mates are not required. The process is rapid and there is enormous production of organisms in a short time and a possibility for successive generations to inherit positive genes from parent organism. In addition, asexual reproduction can occur in various environments. Despite these advantages of asexual reproduction sexual reproduction predominates in the majority of life on Earth because of a number of perks. The benefits of sexual reproduction exceed its drawbacks because it permits genetic mixing, or the uniting of alleles from two parents. Parthenogenesis is a mystery in evolutionary biology since only a small proportion of plant and animal species reproduce asexually. The Red Queen hypothesis, which states that recombination increases the frequency of rare allele combinations that are helpful in fending off attacks from other species or parasites, has been studied in animals that exhibit both sexual and asexual reproduction, including water fleas and New Zealand mud snails. These studies suggest that sexual reproduction may be preferred in changing environments. Recombination, which may distinguish favorable mutations from detrimental ones and boost the efficiency of selection, reduces selective interference, where advantageous mutations fight for fixation in sexual reproduction.
Several hermaphrodite plants and animals employ self-fertilization as a method of reproduction to ensure reproductive success, however, it can cause inbreeding depression owing to harmful mutations. Animals with hermaphroditic sex often avoid self-fertilization. In order to prevent self-fertilization, plants have evolved defense systems (self-incompatibility) including the physical separation of anthers and stigmas. Several species of hermaphrodites still mate once in a while which helps avoid clonal interference and improve adaptability. Humans can experience inbreeding depression as well, which may help to explain why inter-family marriage is stigmatized in society.
How to Be Fit
Species have a broad range of life expectancies, from a few days to possibly indefinite. An organism’s fitness depends on its reproductive tactics, such as fertility, survival, and age of reproduction. Organisms allocate the energy and nutrients they get from their environment to self-maintenance, growth, and reproduction, and there are trade-offs between these functions due to correlated fitness benefits and costs. Reproductive effort is the allocation of nutrients and energy toward reproduction, while the cost of reproduction is the trade-off between reproduction and other functions. Numerous species have provided evidence for a cost of reproduction, since genotypes that devote more resources to reproduction may show reduced survival or development. Reproduction has a cost, according to genetic correlations that have revealed trade-offs between it and fecundity or survival. Fitness in iteroparous species is assessed by lifetime reproductive success, which is estimated by summing the number of births across all ages at which individuals reproduce. The example of an asexual lizard with a three-year lifetime can be used to demonstrate this. In sexually reproducing populations, we can use a life table to calculate the fitness of an allele by using the average values of survivorship and fecundity for individuals carrying that allele.
Natural selection does not act to prolong survival beyond the last age of reproduction, as increasing survival and fecundity at earlier ages has a larger effect on fitness than at later ages. Two major factors are responsible for the evolution of senescence and limited lifespan: mutation accumulation and antagonistic pleiotropy. Mutation accumulation causes mutations that compromise biological functions to reduce fitness less, the later in life they exert these effects, and selection against these mutations is weaker. Antagonistic pleiotropy suggests that many genes affect allocation to reproduction versus self-maintenance and incur a cost of reproduction. Alleles that increase allocation to reproduction early in life reduce function later in life, leading to a negative relationship between early reproduction and both longevity and later reproduction. While both factors can contribute to senescence, many biologists believe that antagonistic pleiotropy is often the more important factor.
The growth rate of a population is affected by ecological conditions, including resources, predation, and disease. The per capita growth rate of a population declines as the population size increases, eventually reaching a stable equilibrium number called the carrying capacity (K), which favors alleles that increase an individual’s ability to compete for limited resources. Species that are adapted to crowded conditions near K are called K-selected, while species that experience rapid, exponential growth and have higher fitness with higher fecundity are called r-selected.
There are different life histories and reproductive strategies in various species. Some species reproduce early and die young, while others delay reproduction and invest in growth and self-maintenance. Species with higher rates of adult survival tend to delay reproduction, while those with low adult mortality may benefit from iteroparity or repeated reproduction. Semelparity, or reproducing only once, may be favored if the probability of survival increases with body mass and if there is an exponential relationship between body mass and reproductive output. Conversely, iteroparity may be advantageous in fluctuating environments or in species with low adult mortality rates.
In species such as humans, albatrosses, and kiwis, there are trade-offs between the number of offspring and their size. Larger offspring require more care and resources from parents, and in some habitats or lifestyles, starting life at a large size greatly enhances the chance of survival. In these species, it may be advantageous to produce only a single, or a few, larger offspring rather than many smaller ones. Additionally, producing too many offspring can lead to decreased survival rates for both parents and offspring, as parents may be unable to adequately feed and care for a large brood. Therefore, the optimal clutch size for a bird, for example, maybe the number of eggs that yields the greatest number of surviving offspring, and this may be fewer than what could be produced given the parent’s resources. Similarly, among plants, larger seeds may be advantageous for species that germinate in the deep shade of closed forests, where the survival and growth of a seedling under adverse conditions are enhanced by the food stored in a large seed’s endosperm.
Sequential hermaphroditism, or the ability to change sex over the course of the life span, can be advantageous in species where reproductive success increases with size to a greater extent in one sex than the other. For example, in species that grow in size throughout reproductive life, a sex change can be beneficial if producing more seeds requires a larger body size, as in the case of squashes and other plants that produce male flowers when small and switch to producing female flowers when larger. In some species of slipper shells and fishes, sex changes occur when individuals reach a certain size or age, with almost all species of sex-changing animals changing sex when they have reached about 70 percent of their maximum size. These sex changes are a result of the costs of reproduction, which affect both males and females and the advantages of larger body size or reproductive output for each sex.
The evolution of life history variation is usually understood in terms of survival and reproduction components of fitness. However, a broader conception of an organism’s life history includes many other aspects of its life, such as dispersal and its use of habitats, food, and other resources. Specialization in ecological niches can be advantageous due to several reasons, including interactions with other species, the advantage of evolving a preference for a safe space, and trade-offs. The plasticity that often underlies broad tolerance can be disadvantageous because it has costs, including the costs of developing an altered phenotype and maintaining the ability to do so. Specialization may also be advantageous due to trade-offs, as a specialist is likely to become more effective or efficient than a generalist, in which performance of any one task is likely to be compromised by the characteristics needed to perform other tasks. Trade-offs in cognitive processing may account for host specialization in some herbivorous insects. Morphological trade-offs have been shown in many organisms, such as flower piercers, which have an unusual hooked bill with which they hold the flower and punch a hole in its base. When the hooked tip of the bill is clipped experimentally, the birds become less efficient at obtaining nectar but become more proficient at eating berries. However, specialization can also be disadvantageous in unpredictable environments, as specialists are more vulnerable to environmental change. Tropical species that already live near their upper thermal limit may be especially endangered by global warming, which could pose a threat to specialized species in general. Overall, understanding the advantages and disadvantages of ecological specialization can shed light on the evolution of life histories and the adaptation of organisms to their environments.
Reference
Futuyma, D. J., & Kirkpatrick, M. (2017). Evolutionary. Evolution (Fourth ed.). pp. 247-292. Sunderland, Massachusetts: Sinauer Associates, Inc.
This post covers chapters 8 & 9 from Futuyma and Kirkpatrick’s book on Evolution (2018). The author of this post is Jacqueline Hoppenreijs.
Evolution in space
Variation in phenotypic traits can reflect variation in environmental factors on a global scale, such as temperature, or on a local scale, such as nutrient or toxicant concentrations. When this phenotypic variation looks like a smooth transition or gradient, we call it a cline. Such ecological variation is a consequence of the selection pressure’s spatial variation and can be seen between and within species, leading to specific patterns of phenotypic traits. An example of such a pattern between species is the occurrence of hairs and cuticles in drought-adjusted plants, that help them retain small amounts of water that they have managed to get a hold of (De Micco & Aronne, 2012). A within-species example is the variation in root depth and structure in individuals of one species that are grown in drier circumstances than others (Lenssen et al., 2004).
Trait patterns are further shaped by exchange between populations of the same species, a process called gene flow that happens through dispersal. Dispersal is different from migration in that it means that an organism ventures out to explore new grounds (or waters) and that it leads to exchange or introduction of genes. Dispersal can be passive (e.g. a seed falls into the water and ends up on a riverbank elsewhere), or active (e.g. an animal starts looking for a more suitable living environment if it runs out of food). There are also forms that could be seen as intermediate, such as plants (Figure 1) that disguise themselves to convince someone else to fix their dispersal for them!
Figure 1. A dung beetle dispersing poop-disguised seeds from the species Ceratocaryum argenteum.
Dispersal can be a bit of a pain, as it requires energy (even if it’s only the effort of growing your seeds in the form of poop) and can be dangerous. It can have big advantages though: by dispersing, you get the opportunity to find a more suitable spot to live and you simultaneously decrease the day-to-day competition with your family members in the place where you all are from. You’re also more likely to reproduce with an individual from a different population, thus decreasing the risk of inbreeding.
So, it’s clear that dispersal can be very profitable, and leads to exchange of genes between populations or establishment of new populations. Gene flow can be measured in multiple ways, that are applicable in discrete (think: migration rate between islands) or continuous (think: migration variance throughout a vast forest) environments, or both. This can be your typical mark-recapture set-up, but more and more often genetics-based methods are used. There are many ways to go about this, but one straightforward approach is to assess genetic difference between populations using their allele frequencies, a variable that then can be combined with the populations’ physical distance to give an idea of gene flow history. Things become less straightforward when there’s local selection involved, that favours one allele over the other and thus (partially) undoes the effect of gene flow. Which of the two processes predominates depends on both local selection and migration rate/variance. If local selection were the dominant process, different populations would have different alleles that fit their environment, and if gene flow were the dominant process, populations would have the exact same allele, also called gene swamping. The two processes often keep each other in somewhat of a balance, and understanding them can help trace past events and processes, and predict future developments (e.g. Love et al., 2023).
Life as an evolutionary biologist is complicated enough with just these two processes going on, but there’s another one that comes into play: genetic drift. This process, that we heard about in blog 3 , is another source of genetic variation and it can be pretty complicated to find out whether it’s this or local adaptation (or both) that cause genetic and phenotypic differences between populations. Mapping of entire genomes makes it possible to look for differences between them, and can offer a solution to this problem. A comparison between the genetic variance in neutral and “under selection” regions in two or more different populations, gives you information on both. As the neutral regions will have a certain amount of variance that’s not a consequence of the environment favouring one allele over the other, this variance is most likely the consequence of genetic drift. The variance in the “under selection” region minus the variance in the neutral regions is than likely to be the result of local adaptation. It’s as simple as that!
Understanding local adaptations in the context of dispersal to new areas can open a whole new can of worms/research questions. One might wonder how other species in the region have adapted to their environment in the past and are now dealing with a new species entering the stage. Ecological research often has a competition-for-resources angle (Simha et al., 2022), but could for example also focus on niche construction (Laland et al., 2016). It’s thus virtually impossible that the entrance of a new species in an area will go unnoticed by the already present species. Researching this, however, requires somewhat of an understanding of how species are defined and come to be, which is our next topic.
Species and speciation
As we know now, species can adapt to new places that they disperse to, and to new circumstances that they’re confronted with. This can ultimately lead to the formation of one or more new species, also called speciation. Before understanding that process, it can be helpful to look at its results: how do we define species as different from each other? The answer is that this is almost always is arbitrary, and the approach can be simple: “they look different”. It gets a bit more complicated with “they can successfully reproduce”, also called the Biological Species Concept (BSC), or quite complicated with the approach “they are the smallest set of organisms that has the same ancestor” which is one of the versions of the Phylogenetic Species Concept (PSC). Why do I write “one of the versions”, you ask? That’s because there’s hardly ever a clear line to draw in biology: think of organisms that don’t sexually reproduce, or different species that hybridise with each other! Besides that, our knowledge and technology keep developing, which means that these concepts are still and probably always will be under debate (see for example Wheeler & Platnick, 2000).
While it’s easy to understand why spatially separated individuals of a species don’t get their genes flowing, there’s a bunch of less well-known mechanisms that prevent gene flow without geographical barriers. These are called Reproductive Isolating Barriers (RIBs), lead to speciation and can be divided in three main categories: pre-mating, pre-zygotic and post-zygotic barriers. Pre-mating RIBs mean that the male and female gamete never get to meet, for example because of a temporal mismatch or because the owners don’t recognise each other’s courting signals. For mating to turn into reproduction, we need a zygote to be formed. If there’s a pre-zygotic barrier such as a mismatch in physique or unsuccessful fertilisation, that means yet another barrier. If all this works and we have a hybrid zygote, it might simply not be able to survive in the environment that its parent organisms are well-adjusted to. Or it might have high mortality in no matter which environment, or is unable to produce offspring, all meaning we have a post-zygotic problem. Post-zygotic barriers are often caused by mismatches between the genes of the respective parents and are often found on the site of the heterogametic sex, i.e. the counterparts of humans that have XY instead of XX chromosomes. Different forms of reproductive isolation (RI) aren’t equally important, don’t necessarily occur equally fast in time and affect each other. These complications mean that experiments and technology in the lab and the field are incredibly important for our understanding of what’s (been) going on.
If we take a step back, we can at least say that evolutionary biologists have a pretty good understanding of what might be happening in individuals and their populations, and what processes and mechanisms can lead to the origination of a new species. There are several causes that can drive the processes that lead to speciation:
Ecological: two populations of one species adapt to different environments through natural selection;
Genetic conflict: a certain allele becomes very abundant but has negative effects on fertility, and another mutation, that repairs these effects, is incompatible with the “original” genetical set-up;
Sexual selection: sex A prefers certain visual, audio or other traits in sex B over other trait expressions, leading to certain varieties of certain traits always reproducing with each other instead of with other varieties in the species;
Reinforcement of RI: hybrids of two populations have lower survival or fertility meaning that they’ll be less successful reproducing than individuals from the two populations within themselves;
Polyploidy: genome-duplication can result in a new species within one generation when tetraploid individuals are unable to reproduce successfully with individuals from the original, diploid populations;
Hybrid speciation: the hybrid(s) of two “parent” species become genetically incompatible with them;
Genetic drift: a population that becomes fixed for a chromosomal rearrangement, a consequence of genetic drift, may no longer be compatible with the rest of the species.
The latter, speciation through genetic drift, occurs especially in small or highly-fluctuating populations. It is an important pillar under the founder effect: the start of a new population by a few individuals. If these few individuals have undergone changes due to genetic drift and get isolated from the rest of the population, they can come to form a new species if their genetic make-up is incompatible with the population of origin. This can be especially interesting at the edges of species ranges (Figure 2a), and there are descriptions of how this process can occur multiple times in a row, leading to a “wave” of changes through mutation (Figure 2b). If this leads to allele frequencies becoming relatively and unexpectedly high, this is called genetic “surfing” (Excoffier et al., 2009).
Figure 2. Allele frequencies a) during range expansion and b) during serial range expansion events. Derived from (Peischl et al., 2016).
While the founder effect is not fully understood or 100% supported by the literature, it’s clear that the geographical aspect that underlies it plays a role in many speciation events. If gene flow is completely blocked off by a geographical barrier, we speak of allopatric speciation. Note that the degree of geographic isolation depends on a geographic barrier, not on geographic distance, and that the degree to which it is an actual barrier also depends on the dispersal capacities of the species in question. Barriers can have their source in the environment, e.g. a new river divides two grasslands in two, or in a species itself, e.g. a few individuals manage to leave the mainland to start a population on an uninhabited island. Both can lead to allopatric speciation, but neither of these two processes has to be irreversible, as the river can fall dry again or the island species might expand their range to the mainland. In both situations, the populations of the mainland and the island can become sympatric. In such cases, we speak of secondary contact between the populations where, depending on the degree of reproductive isolation, gene flow is possible.
Sympatric speciation and the intermediate process, parapatric speciation, still have some gene flow going on. The former doesn’t depend on geographical barriers, but can depend on factors such as timing (e.g. flowering plants), local variation (e.g. soil characteristics) and behaviour (e.g. mating preferences). Sympatric speciation can reinforce itself if the allele in question is associated to another specific allele on another gene, which is rare but can occur when there’s a trait variety that works on both ecological divergence and reproductive isolation. Called a speciation trait, this trait can for example cause individuals with a specific feeding preference to only reproduce with each other. Other non-random mating patterns can lead to parapatric speciation. This leads to subpopulations continuing to interbreed with each other while getting more reproductively isolated from each other.
While the patterns of speciation are not easily understood and mapping them can become easier in collaboration with fields such as paleoarchaeology and geology, there is a lot of variation within the genome that’s left to be unravelled. The omics side of evolutionary biology can further the understanding of linkage equilibria, self-reinforcing processes and speciation genes and regions, to be able to understand what has happened in the past. Given the current pace of ecosystem degradation and destruction, we need this understanding of evolution to predict and, where possible, adapt to the future.
References
De Micco, V., & Aronne, G. (2012). Morpho-Anatomical Traits for Plant Adaptation to Drought. In R. Aroca (Ed.), Plant Responses to Drought Stress (pp. 37–61). Springer. https://doi.org/10.1007/978-3-642-32653-0_2
Excoffier, L., Foll, M., & Petit, R. J. (2009). Genetic consequences of range expansions. Annual Review of Ecology, Evolution, and Systematics, 40, 481–501. https://doi.org/10.1146/annurev.ecolsys.39.110707.173414
Futuyma, D., & Kirkpatrick, M. (2018). Evolution (4th ed.). Oxford University Press.
Laland, K., Matthews, B., & Feldman, M. W. (2016). An introduction to niche construction theory. Evolutionary Ecology, 30(2), 191–202. https://doi.org/10.1007/s10682-016-9821-z
Lenssen, J. P. M., Van Kleunen, M., Fischer, M., & De Kroon, H. (2004). Local adaptation of the clonal plant Ranunculus reptans to flooding along a small-scale gradient. Journal of Ecology, 92(4), 696–706. https://doi.org/10.1111/j.0022-0477.2004.00895.x
Love, S. J., Schweitzer, J. A., & Bailey, J. K. (2023). Climate‑driven convergent evolution in riparian ecosystems on sky islands. Scientific Reports, 1–9. https://doi.org/10.1038/s41598-023-29564-2
Peischl, S., Dupanloup, I., Bosshard, L., & Excoffier, L. (2016). Genetic surfing in human populations: from genes to genomes. Current Opinion in Genetics and Development, 41, 53–61. https://doi.org/10.1016/j.gde.2016.08.003
Simha, A., Pardo-De la Hoz, C. J., & Carley, L. N. (2022). Moving beyond the “Diversity Paradox”: The Limitations of Competition-Based Frameworks in Understanding Species Diversity. American Naturalist, 200(1), 89–100. https://doi.org/10.1086/720002
Wheeler, Q. D., & Platnick, N. I. (2000). The Phylogenetic Species Concept (sensu Wheeler and Platnick). In Q. D. Wheeler & R. Meier (Eds.), Species concepts and phylogenetic theory – a debate (pp. 55–68). Columbia University Press.
This post covers chapters 6 & 7 from Futuyma & Kirkpatrick’s book on Evolution (2018). The author ofthis post is Jeff Marker.
Phenotypic Evolution
One major key to understanding how evolution works is based on the genetical theory related to inheritance, selection, and fitness. Through phenotypic evolution, we will now examine the quantitative traits produced by those genetic mechanisms, how the environment affects such traits, and how those traits drive evolution. Simply put, a quantitative trait is a species phenotype that is measurable in a continuous way. Quantitative traits are controlled and affected by a combination of many different genes and the environment. For example, blood pressure in humans is a product of environmental factors such as available diet in combination with genes that control a complex system of fluids, hormones, and salts in the body. Quantitative traits vary between species, but, like most phenotypes, they tend to follow a normal distribution over the entire population.
Natural selection acts on quantitative traits though the mechanism of fitness function where a specific trait of a species acts on its ability to survive or reproduce. There are three different types of selection associated with quantitative traits and fitness function: directional, stabilizing, and disruptive. First, directional selection favors either an increase or a decrease in a trait’s mean in the population, favoring individuals at one extreme of the phenotypic distribution. For example, the females of the spider Nephila clavipes (Figure 1) are selected for large body sizes as they are able to construct larger webs that capture more prey and can produce more offspring. Second, stabilizing selection is seen in species that have trait values near the population mean with intermediate phenotypes. Species of the orb-weaving spider, Argiope aurantia, capture more prey with intermediate sizes of web decoration versus other individuals with very large or very small decoration. Finally, disruptive selection is a case where both extremes of a trait are advantageous and the intermediate individuals tend to be less common. Females of the spider Gasteracantha cancriformis, prefer males with either very high levels of spines on the abdomens or low levels of spines on the abdomen and often reject those with intermediate levels of spines leading to a bimodal distribution of the number of spines on male abdomens within this species population.
Each of these types of selection is a piece of the driving force of evolution. Previously we have learned that as natural selection acts on a trait and that trait is successfully passed on to subsequent generations, then evolution of the species in question results. But the importance of quantitative traits lies in our ability to measure them and use them to make meaningful measurements about the rate of evolution. Through directional selection we can measure trait means at the start of a generation and the means of the individuals that survive and reproduce. This helps us understand the heritability, or the strength of inheritance, within a population of a species and ultimately we can calculate the rate of evolution based on that. Heritability is one piece of the overall phenotypic variance in a population. The other two sources of that variance is determined by environmental variance, such as climate or available nutrition sources, and gene dominance and epistasis. Gene dominance is where one allele expresses its phenotype over another and in epistasis a gene modifies the expression of another gene.. Natural selection acts on the existing, or standing, genetic variation within a population favoring certain traits or combinations of traits that increase an organism’s fitness. In contrast, populations with low levels of standing genetic variation may be less able to respond to changing environmental conditions, making them more vulnerable to extinction. Selection pressure, environment, and gene function all work simultaneously to determine a specie’s phenotypic traits
Humans have been using artificial selection for thousands of years to breed and improve a variety of plants and animals to aid in agricultural, industrial, and cultural advancement of our species. As a result of artificial selection, we have revealed a handful of general conclusions about phenotypic and trait based selection:
When a trait is selected, it will evolve
Traits can evolve beyond the variation that was present in the original population
Small populations evolve slower than large populations
Trait selection can cause other traits to be negatively affected
There exists a constant and strong trade-off regarding trait selection and evolution. This idea can often be boiled down to the phrase “there are no free rides in nature.” Traits are correlated. A common example is that with spiders. Large spiders tend to build larger webs while smaller spiders build smaller webs. These correlations between two (out of many) traits contribute to the overall fitness of the spiders and affect their ability to catch prey, mate, and provide specific functions within their environments. Some evolutionary trade-offs can help increase species fitness while other trade-offs balance each other out or may actually create a negative genetic correlation. For example, females in the larger species of spiders have higher fecundity and produce more eggs than small species. One trade-off is that these larger species have longer development times that often result in higher mortality rates.
Some species change their phenotypic expression as a result of their environmental conditions. This is called phenotypic plasticity. Plasticity can be either irreversible or reversible and may affect morphology, physiology, or behavior. Some web-weaving spiders will change the location, size, and shape of their webs in response to weather conditions, prey availability, or to better protect themselves from predators. These types of responses are not because of genetic changes, but rather a direct result of the surrounding environment.
Due to significant advances in DNA technology, we can now determine exactly what portions of a genome affect quantitative traits. Some quantitative traits are affected by a single locus with alleles that have large and significant effects while other quantitative traits are coded for with multiple genes. Understanding exactly which genes, alleles and nucleotides affect traits is imperative to better understand the mechanisms of evolution. Further, finding the quantitative trait loci can assist researchers in finding new therapies for infections that have generated resistance to current drugs.
Genetic drift
Chance is always a factor in evolution. We often like to idealize genomics, natural selection, and evolution as being this uncaring machine that just works. Traits are selected, species reproduce, populations evolve, or populations do not evolve. We learned in previous chapters about trait selection and reproduction and how this drives species evolution. However, what if this was all just a coin flip? What if the color of your hair, the size of a snail’s shell, or the number of leaves on a sunflower plant were all just due to random chance? Randomness and chance are both important pieces of the concept of genetic drift. Genetic drift is a random process causing changes in the frequency of traits within a population. Yes, both natural selection and genetic drift drive species evolution by varying the gene frequency within a population over some defined time. However, it is imperative to point out that genetic drift fluctuates due to chance events whereas natural selection is a process of species adaptation driven by environmental changes. Something that would be difficult to find here in Northern Europe but is nonetheless an accurate example of genetic drift in action is the unfortunate trait of lactose intolerance in humans. Genetic drift was likely an important factor in the initial spread of lactose tolerance. A single individual with the gene for lactose tolerance may, due to chance events, have had many descendants and thus increased that gene frequency over time within certain populations of humans. Although after initial rounds of genetic drift of lactose tolerance in humans, it is likely that positive selection played a role in its proliferation in Northern Europe.
Genetic drift can be particularly important in small populations, where chance events can have a larger impact. As a result, genetic drift can reduce genetic variation within a population, and can even lead to the loss of certain alleles. Other alleles can get fixated, meaning that they become the only allele for one locus present in the population.
There are five central features of genetic drift:
1. Genetic drift is impartial. Allele frequency in a population is just as likely to go up as it is to go down. Remember, genetic drift is due to chance, so alleles that move via genetic drift do so at random. Natural selection on the other hand can favor one allele over another as they are often chosen as a way to increase fitness.
2. Smaller populations are more susceptible to genetic drift. This causes larger fluctuations in allele frequency in smaller populations vs larger ones as the allele shift in larger populations tends to average out based on the larger number of species.
3. Genetic drift causes an overall loss in genetic variation in species populations. Once an allele is fixed within the genetics of a population, it is then the only one that will be passed on to subsequent generations.
4. Genetic drift changes the genomes of identical populations leading to more genetic variation on a larger scale.
5. Alleles can become fixed in a population without any assistance from natural selection.
Genealogy mapping is used to determine the positions of genes on a chromosome and their inheritance patterns. This mapping is used to understand the relationship between genetic variation and the physical traits that they are associated with. We know that different genes have different genealogies. Genes on a chromosome can be viewed similarly to human beings in a country: they have different common ancestors. Those common ancestors may have originated from many different places at many different times. For example, mitochondrial DNA (mtDNA) in humans can be traced back about 125,000 years ago to one of our common ancestors. In fact, mtDNA in all eukaryotic species can be mapped and traced back to a single common ancestor that lived about 2 billion years ago.
Genetic drift is always at work. However, the strength of that genetic drift is dependent on the effective population size (Ne). Ne is the number of individuals in a that would lose genetic variation at the same rate as the real population. Ne is influenced not only by the number of individuals but also the number that are sexually mature, the strength of random mating between individuals, and the sex ratio. In general, larger populations are less susceptible to drift than smaller populations, but only an infinitely large population would never experience genetic drift. Two types of genetic drift that work strongly on small populations are the bottleneck effect and the founder effect. The bottleneck effect occurs when a population undergoes a dramatic reduction in size, which can lead to a loss of genetic variation due to the random elimination of alleles. The founder effect occurs when a new population is established by a small group of individuals, which can lead to the loss of genetic variation from the original population. For example, the lake whitefish (Coregonus clupeaformis) in the Great Lake region of the North America have suffered major declines in genetic diversity due to intense fishing pressure over the last 100 years. As such, these small populations have been found to have very low genetic diversity likely as a result of being founded by a very small number of individuals (Graham et al., 2022; Stott et al., 2013). These events can be quite damaging to populations as low genetic diversity can lead to species’ being unable to adapt to changing environments.
Coregonus clupeaformis (National Oceanic and Atmospheric Association, USA)
Genetic drift and natural selection work together to move advantageous mutations through a population. Sometimes they work together to increase the mutation frequency, but occasionally, especially in smaller populations, genetic drift can cause an advantageous mutation to decrease or even be completely removed from a population. Drift can also be responsible for harmful mutations to spread and eventually become fixed. Again, this can be common in small populations that are bottle-necked sometime during their history or in populations with significant inbreeding. Inbreeding load is the weakening of fitness in a small population where harmful genetic mutations become fixed in the genome. Here, inbreeding load is different from inbreeding depression where closely related parents produce offspring with reduced fitness. Florida panthers (Puma concolor coryi) in the southern reaches of Florida have declined into the hundreds and as a result, significant inbreeding load has caused this species to have many health and physical problems including heart defects and lower reproductive success. Unfortunately, genetic drift also causes highly beneficial mutations to become completely lost in a population. This essentially acts as an “evolutionary speed limit” on how quickly species adapt to environmental changes.
Gene adaptation and local adaptation are related mechanisms in evolution referring to different ways in which populations adapt to changing environmental conditions. Gene adaptation refers to the evolution of specific genetic variants. This typically happens through natural selection where beneficial and harmful alleles increase and decrease in frequency, respectively. Local adaptation refers to the evolution of populations that are adapted to their local environment. Local adaptation can involve both genetic and non-genetic factors such as changes in morphology and behavior.
References
Graham, C. F., Boreham, D. R., Manzon, R. G., Wilson, J. Y., & Somers, C. M. (2022). Population structure of lake whitefish (Coregonus clupeaformis) from the Mississippian lineage in North America. FACETS, 7(1), 853-874.
Stott, W., Ebener, M. P., Mohr, L., Hartman, T., Johnson, J., & Roseman, E. F. (2013). Spatial and temporal genetic diversity of lake whitefish (Coregonus clupeaformis (Mitchill)) from Lake Huron and Lake Erie. Adv Limnol, 64, 205-222.
RivEM – Karlstad University
… is a research group associated to the subject of Biology and the Department of Environmental and Life Sciences at Karlstad University, Sweden. We conduct both basic and applied research on human impact on river ecosystems, and how this impact can be minimized.
In Swedish the research group is called Naturresurs Rinnande vatten (Acronym NRRV, hence the url nrrv.se).